Pharmaceutical Sciences. 31(4):452-463.
doi: 10.34172/PS.025.41060
Research Article
Bergenin Synergistically Enhances the Efficacy of Cisplatin and 5-FU: Strategy to Enhance Chemotherapy in Breast and Colorectal Cancer - An In Vitro Study
Priyesh Kumar Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing, 1 
Khairah Ansari Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing, 1
Vaibhavi Srivastava Formal analysis, Investigation, Writing – review & editing, 1
Ann Maria Joseph Formal analysis, Investigation, Writing – review & editing, 1
Nilam Parmar Methodology, 2
Devendrasinh Jhala Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review & editing, 1, *
Author information:
1Department of Zoology, University School of Sciences, Gujarat University, Ahmedabad, Gujarat, India 380009
2Department of Life Sciences, University School of Sciences, Gujarat University, Ahmedabad, Gujarat, India 380009
Abstract
Background:
The cytotoxic and synergistic efficacy of bergenin, a natural compound, against MCF-7 breast cancer and HCT 116 colorectal cancer cell lines was investigated. Bergenin, combined with cisplatin (Cis) and 5-fluorouracil (5-FU), can enhance the efficacy of chemotherapy and be ultimately useful for dose reduction of chemotherapeutic agents.
Methods:
The in vitro study involved treating MCF-7 and HCT 116 cells with varying doses of bergenin, Cis, and 5-FU for 24 and 48 hours. The cytotoxic effects were measured using MTT assay, while synergistic activity was evaluated using CompuSyn. Fluorescent staining, colony forming, wound healing, cell cycle and apoptosis assays were performed to support the findings. Statistical significance less than 0.05 was considered and assessed using ANOVA (one-way or two-way) and Tukey’s test.
Results:
Bergenin showed a dose- and time-dependent inhibition of cell proliferation in both MCF-7 and HCT 116 cells, at 24 and 48 hours, respectively. Combining bergenin with Cis or 5-FU led to significant reductions (P<0.0001) in their IC50 values. Multiple lines of evidence, including fluorescent staining, colony forming, wound healing, cell cycle and apoptosis assays, corroborated the statistically significant cytotoxic and synergistic effects of bergenin compared to control.
Conclusion:
This study underscores bergenin’s potential as a therapeutic agent in cancer treatment. Bergenin inhibits cell growth, induces apoptosis, and enhances the efficacy of Cis and 5-FU, suggesting its potential as an addition to existing treatment regimens. This necessitates further research, elucidating the clinical applications of bergenin in cancer therapy, which could improve treatment outcomes and reduce adverse effects associated with high doses of chemotherapeutic agents.
Keywords: 5-Fluorouracil, Anticancer, Bergenin, Breast and colorectal cancer, Cisplatin, Synergism
Copyright and License Information
© 2025 The Author(s).
This is an open access article and applies the Creative Commons Attribution Non-Commercial License (
http://creativecommons.org/licenses/by-nc/4.0/). Non-commercial uses of the work are permitted, provided the original work is properly cited.
Funding Statement
This study was self-funded by the authors and received no external financial support from any funding organization.
Introduction
Breast cancer, the most frequently diagnosed cancer among women globally, and colorectal cancer, which affects both men and women, pose significant public health challenges due to their widespread prevalence and profound impact on patient outcomes. Despite advancements in screening techniques and treatment modalities, breast cancer remains a leading cause of cancer-related morbidity and mortality among women worldwide.1 Similarly, colorectal cancer, originating in the colon or rectum, ranks among the most prevalent and deadliest cancers worldwide.1 The multifactorial nature of both breast cancer and colorectal cancer underscores the complexity of these diseases, involving genetic, environmental, and lifestyle factors.
Bergenin, a natural compound, exhibits strong antitumor effects across multiple cancer types, including cervical carcinoma, colorectal cancer, oral squamous cell carcinoma, bladder cancer, and non-small cell lung cancer. It effectively reduces cancer cell viability and colony formation and inhibits tumor growth in both in vitro and in vivo models by targeting critical signaling pathways and proteins. In 2022, Sultana et al2 demonstrated that the aqueous extract of E. agallocha inhibits the growth of SiHa cervical cancer cells by inducing autophagy, apoptosis, mitophagy, and G2/M phase arrest, with bergenin identified as the key anti-cancer agent. In cervical carcinoma, bergenin specifically targets angiogenic proteins such as galectin-3 and MMP-9.3 In colorectal cancer, it downregulates the Akt/GSK3β pathway and promotes Mcl-1 degradation.4 In oral squamous cell carcinoma, bergenin upregulates PTEN and inhibits the AKT/HK2 axis, thereby suppressing glucose metabolism.5 Moreover, bergenin activates the PPARγ/PTEN/Akt pathway in bladder cancer and reduces survivin expression in non-small cell lung cancer by inhibiting the Akt/Wee1/CDK1 pathway.6 Through its ability to induce apoptosis, autophagy, mitophagy, and cell cycle arrest, bergenin emerges as a promising candidate for anticancer therapy across a wide range of malignancies.7
Although cisplatin (Cis) and 5-fluorouracil (5-FU) have demonstrated efficacy in combating breast and colorectal cancer respectively, their utility is often hindered by the occurrence of adverse side effects. Cis, a platinum-based chemotherapy drug widely used in various cancer types including breast and colorectal cancer, can induce severe adverse effects such as nephrotoxicity, neurotoxicity, gastrointestinal disturbances, and bone marrow suppression.8 Similarly, 5-FU, a cornerstone in the treatment of colorectal cancer, is associated with gastrointestinal issues such as nausea, vomiting, diarrhea, mucositis, as well as myelosuppression, fatigue, and increased susceptibility to infections.9 In response to these challenges, researchers are exploring innovative approaches to cancer treatment, including the investigation of adjunctive therapies. The study addresses a noted gap in the literature regarding the potential synergistic effects of combining the natural compound bergenin with standard chemotherapeutic agents, specifically Cis and 5-FU, for treating breast (MCF-7 cell line) and colorectal (HCT 116 cell line) cancer. Despite bergenin’s known benefits, studies on its synergistic effects with chemotherapy are lacking. Therefore, the aim of this study is to enhance the effectiveness of Cis and 5-FU by utilizing bergenin, potentially allowing for lower dosages and a subsequent reduction in associated side effects. The underlying hypothesis is that combining bergenin with Cis or 5-FU will produce a synergistic effect. This synergy is expected to enhance the cytotoxic efficacy of the chemotherapy drugs, potentially allowing for reduced dosages, which could consequently minimize associated adverse side effects and improve treatment tolerability.
Methods
Cell culture maintenance and experimental groups
The breast cancer cell line MCF-7 and colorectal cancer cell line HCT 116 were obtained from National Centre for Cell Science (NCCS), Pune, Maharashtra, India. MCF-7 cells were grown in RPMI 1640, and HCT 116 cells in DMEM medium, both supplemented with 10% fetal bovine serum (FBS) and a 1 % pen/strep solution (100 units/mL penicillin and 100 µg/mL streptomycin). The cells were incubated with humidified 5% CO2 at 37 °C.MCF-7 cells were subjected to 35 μM bergenin, 14 μM Cis, and a combination of 20 μM bergenin with 5 μM Cis for 24 hours. Likewise, HCT 116 cells were treated with 40 μM bergenin, 23 μM 5-FU, and a combination of 10 μM bergenin with 10 μM 5-FU for the same time period as shown in Table 1.
Table 1.
Distinct groups, treatments and concentration based on analysis of combination study
|
Groups
|
Cell line
|
Treatment*
|
Concentration (µM)
|
| I |
MCF-7 |
Untreated |
- |
| II |
Bergenin |
35 µM |
| III |
Cis |
14 µM |
| IV |
Bergenin + Cis |
20 µM + 5 µM |
| V |
HCT 116 |
Untreated |
- |
| VI |
Bergenin |
40 µM |
| VII |
5-FU |
23 µM |
| VIII |
Bergenin + 5-FU |
10 µM + 10 µM |
* < 0.1% DMSO was maintained for all the experimental groups.
In vitro cytotoxicity assay using 3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay
The in vitro anticancer activity of bergenin was assessed using the MTT assay on MCF-7 and HCT 116 cell lines, as previously outlined by Mosmann.10 MCF-7 and HCT 116 cells were seeded in 96-well plates (104 cells/well in 200 μL of media) and allowed to adhere for 24 hours. MCF-7 cells were treated with bergenin and Cis, and HCT 116 cells with bergenin and 5-FU, each at concentrations of 5-50 μM. After a 24- and 48-hours treatment with the test compounds, the culture medium was removed, and the cells were rinsed with PBS. Subsequently, 50 µL of MTT reagent (5 mg/mL in PBS) was added into each well and incubated for 4 hours at 37 °C in a humidified incubator with 5% CO2, shielded from light, to facilitate the formation of formazan crystals. Afterward, MTT was removed, and 100 μL of DMSO was added to dissolve the formazan crystals. The plate was further incubated for 30 minutes, and the absorbance was measured at 570 nm using the BioTek Epoch Microplate Spectrophotometer (Agilent, USA). Cell viability (%) was calculated using the given formula:
Where, At is the absorbance of test samples and Ac is the absorbance of untreated control.
Analysis of combination and dose reduction index
The combination index (CI) is a graphical measure of drug interactions, indicating synergism, additivity, or antagonism based on whether the CI is less than, equal to, or greater than 1, respectively. For combination studies, bergenin was tested at concentrations of 10, 20, and 30 μM in combination with varying concentrations of Cis (2.5-20 μM) and 5-FU (5-40 μM) for MCF-7 and HCT 116 cell lines, respectively for 24 hours. Combination analysis was conducted using CompuSyn 1.011 based on Chou-Talalay method.12 To observe the gradual decrease in cell viability among the combination groups, the percentage cell viability of different combinations was plotted against varying concentrations of Cis and 5-FU. Additionally, the CI was calculated at different combination concentrations to identify specific doses that exhibit high synergism with a 50% effect. Normalized isobologram, a dose-oriented graph based on the CI equation at non-constant ratios of different combination groups was constructed. Finally, the dose reduction index (DRI) was calculated at 50% effect level for lowest CI. This dose-reduction defines the combination doses for each positive control (i.e., Cis and 5-FU) with bergenin.
Fluorescent staining
For examining induction of apoptosis using fluorescence staining, MCF-7 and HCT 116 cells were seeded in 12-well plates (1 × 105 cells/well in 1 mL of media) and allowed to adhere for 24 hours. Following exposure to test compounds for 24 hours, the culture medium was removed, and cells were washed with PBS before fixation with 4% paraformaldehyde for 10-12 minutes. The cells were treated with respective fluorescence stains, rinsed with PBS, and then examined using a ZOE fluorescent cell imager (Bio-Rad) at 20 × magnification.
Dual (AO/EtBr) fluorescent staining is employed to detect cellular alterations associated with apoptosis and to distinguish cells at various stages of apoptosis.13,14 The cells were stained with AO/EtBr (100 μg/mL) solution for 10 minutes, followed by another PBS wash.
Hoechst 33342 effectively labelled the nuclei of both viable and non-viable cells, while propidium iodide (PI) specifically labelled the nuclei of non-viable cells.15 The cells were stained with PI (10 μg/mL) solution for 10 minutes, followed by a PBS wash and then stained with Hoechst 33342 (10 µg/mL) solution for 10 minutes.
DAPI, a fluorescent dye that specifically binds to DNA, is commonly used to evaluate DNA fragmentation during apoptosis. Staining was conducted based on the method outlined by Atale et al16 using a 25 μg/mL DAPI solution for 10 minutes.
Colony forming assay
MCF-7 and HCT 116 cells were seeded at a density of 3 × 105 cells per well in 6-well plates, allowed to adhere for 24 hours, and then treated with the test compounds for an additional 24 hours. After the treatment duration, cells were trypsinized and 1 × 103 cells were seeded into each well and maintained in a 5% CO2 environment at 37 °C for two weeks, with media refreshed every 72 hours. Following this, colonies were immobilized using 500 µL/well chilled aceto-methanol solution (1:3 ratio) for 20-30 minutes and then stained with 0.5% crystal violet. Subsequent to capturing images, analysis was conducted using ImageJ software (version 1.54f). The survival fraction (percentage of colonies) was calculated as per the Kabakov et al17 using the given formula:
Where, Ct is the number of colonies formed in treatment groups, Cc is the number of colonies formed in untreated control group.
Wound healing assay
To explore the effect of test compounds on cancer cell migration, the wound healing assay, also known as the scratch assay18 was performed. MCF-7 and HCT 116 cells were seeded in 6-well plates at a density of 3 × 105 cells/well. Upon reaching 80% confluence, a gentle scratch was made across the center of each well using a sterile 200 µL pipette tip. Following the removal of detached cells by washing with PBS, the cells underwent treatment with various test compounds for 24, 48, and 72 hours. Throughout the exposure period, the cells were cultured in medium supplemented with 5% serum. The closure of the wound was monitored at 24-hours intervals from 0 to 72 hours. Photographs were captured using a Zeiss Primovert inverted microscope at 10 × magnification, and the images were analyzed using ImageJ software (version 1.54f).
Cell cycle analysis
The cell cycle was evaluated using a DNA-based flow cytometry method, following the protocol outlined by Chung et al.19 MCF-7 and HCT 116 cells were cultured in 6-well plates at a density of 3 × 105 cells/well and allowed to adhere for 24 hours. Following this, cells were treated with test compounds for an additional 24 hours. Cell harvesting was conducted by incubating them with 300 µL of hypotonic staining solution, comprising 100 mg sodium citrate, 30 µL Triton-X 100, 5 mg PI, and 0.002 mg Ribonuclease A in 100 mL distilled water, for 15 minutes. Analysis of the samples was conducted using the BD FACSAriaTM Fusion Flow Cytometer (BD Biosciences, USA). Data analysis and results were obtained using FCS ExpressTM 7 (Dotmatics, USA).
Cell apoptosis analysis
Early apoptosis involves various morphological and biochemical changes, such as membrane blebbing and the externalization of phosphatidylserine (PS) on the cell surface, which can be detected through Annexin V binding. Through the application of flow cytometric analysis and Annexin V coupled with FITC, apoptotic cells can be effectively identified. Apoptosis in cells was evaluated using the BD PharmingenTM FITC Annexin V Apoptosis Detection Kit I, following the method described by Jin et al., with slight modifications.20 MCF-7 and HCT 116 cells were seeded in 6-well plates at a density of 3 × 105 cells/well and treated with different concentrations of the test compounds for 24 hours. After washing with cold PBS, cells were resuspended in 1 × binding buffer, and a mix of 5 µL of FITC Annexin V and 5 µL of PI was added. The mixture was then incubated for 15 mins at room temperature in the dark before analysis using the BD FACSAriaTM Fusion Flow Cytometer (BD Biosciences, USA). Data analysis and results were obtained using Floreada (https://floreada.io, accessed 26th March 2024).
Statistical analysis
All experiments were conducted separately in triplicate and are expressed as mean ± SD. The statistical significance was evaluated using one-way or two-way analysis of variance (ANOVA), and the individual comparison was obtained by Tukey’s multiple comparison test using GraphPad Prism (version 9.4.0). P < 0.05 was considered to indicate a statistically significant difference.
Results
In vitro cytotoxicity assay using 3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay
The results revealed a dose- and time-dependent inhibition of cell growth in both the MCF-7 and HCT 116 cell line, as depicted in Figure 1. In MCF-7 cells, bergenin demonstrated an IC50 of 34.16 ± 0.57 μM and 20.60 ± 0.60 μM after 24 and 48 hours, respectively as shown in Table 2. Comparatively, the positive control Cis exhibited an IC50 of 14.43 ± 0.37 μM and 7.86 ± 0.74 μM at 24 and 48 hours. In the case of HCT 116 cells, bergenin displayed an IC50 of 40.93 ± 0.40 μM and 25.50 ± 0.69 μM after 24 and 48 hours, while the positive control 5-FU demonstrated an IC50 of 22.87 ± 0.46 μM and 15.04 ± 0.98 μM at 24 and 48 hours, respectively (Table 2). These findings underscore the cytotoxic efficacy of bergenin against both breast and colorectal cancer cell lines.
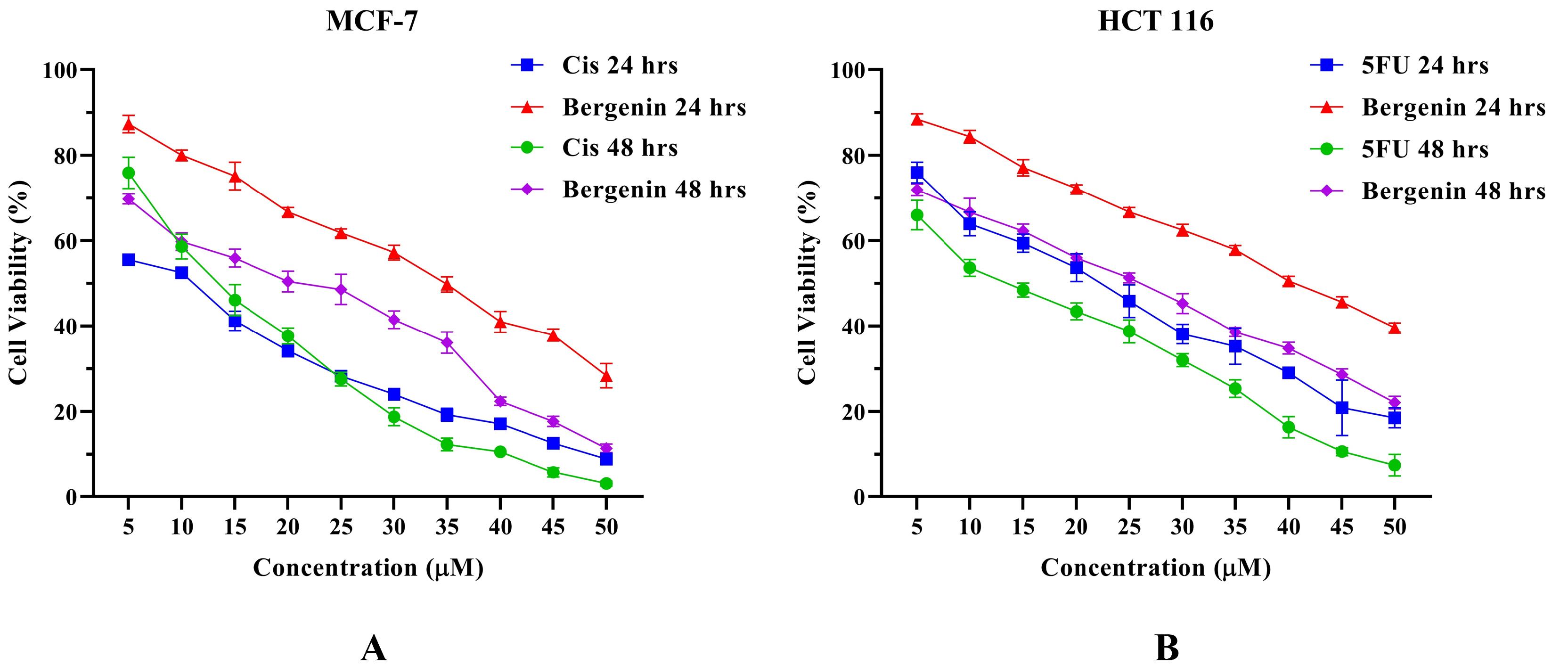
Figure 1.
Cell viability (%) of (A) MCF-7 cells and (B) HCT 116 cells treated with Bergenin compared to positive control Cis and 5-FU, respectively, at different concentrations for 24 and 48 hrs. Values are expressed in mean ± SD
.
Cell viability (%) of (A) MCF-7 cells and (B) HCT 116 cells treated with Bergenin compared to positive control Cis and 5-FU, respectively, at different concentrations for 24 and 48 hrs. Values are expressed in mean ± SD
Table 2.
IC50 of bergenin for MCF-7 and HCT 116 cells compared to positive control Cis and 5-FU, respectively for 24 and 48 hours
|
IC50 in µM
|
MCF-7
|
HCT 116
|
|
24 hours
|
48 hours
|
24 hours
|
48 hours
|
| Bergenin |
34.16 ± 0.57 |
20.60 ± 0.60 |
40.93 ± 0.40 |
25.50 ± 0.69 |
| Cis |
14.43 ± 0.37 |
7.86 ± 0.74 |
- |
- |
| 5-FU |
- |
- |
22.87 ± 0.46 |
15.04 ± 0.98 |
Values are expressed as mean ± SD.
Analysis of combination and dose reduction index
The combination strategy involving bergenin along with the drug molecules used in the treatment of breast and colon cancer i.e., Cis and 5-FU respectively (positive controls) demonstrated a synergistic effect in inhibiting the growth and proliferation of both MCF-7 and HCT 116 cells (Supplementary file 1, Figures S1 and S2). After evaluation of CI, it was observed that among the 15 data points representing various combinations of bergenin with Cis, two of them exhibited antagonism (CI > 1), two were nearly additive, while the rest demonstrated synergism (CI < 1). Conversely, when combined with 5-FU, only one data point indicated antagonism, with all other points displaying synergistic effects Figure S3 (Supplementary file 1). Notably, both bergenin + Cis and bergenin + 5-FU combinations exhibited synergism at a 50% effect level. These groups revealed the dose dependent inhibitory effects.
Analysis through the DRI revealed significant reductions in the IC50 values for the combination groups. DRI > 1 indicates favorable and DRI < 1 indicates unfavorable dose-reduction. For the combination of bergenin with Cis, the DRI indicated a substantial decrease in IC50 values. Specifically, the combination of 10, 20, and 30 µM bergenin with Cis resulted in dose reductions of 1.45, 4.01, and 19.13-fold (P < 0.0001), respectively, while the combination of 10, 20, and 30 µM bergenin with 5-FU resulted in dose reductions of 1.93, 3.05 and 22.29-fold (P < 0.0001) as highlighted in Table 3. This suggests a synergistic enhancement of cytotoxic effects when bergenin is combined with Cis and 5-FU, emphasizing the potential for dose optimization in cancer treatment.
Table 3.
IC50 of positive control Cis and 5-FU alone and in combination with bergenin (10, 20 and 30 µM) and Drug reduction indices of combination groups
|
Combination group
|
IC50 in uM
|
DRI (Dose reduction index)
|
| Cis alone |
15.38 ± 0.31 |
|
| 10 μM Bergenin + Cis |
10.64 ± 0.17**** |
1.45 |
| 20 μM Bergenin + Cis |
3.84 ± 0.29**** |
4.01 |
| 30 μM Bergenin + Cis |
0.80 ± 0.06**** |
19.13 |
| 5-FU alone |
25.17 ± 0.01 |
|
| 10 μM Bergenin + 5-FU |
13.02 ± 0.41**** |
1.93 |
| 20 μM Bergenin + 5-FU |
8.23 ± 0.97**** |
3.05 |
| 30 μM Bergenin + 5-FU |
1.13 ± 0.22**** |
22.29 |
Results are expressed as mean ± SD. Statistical comparisons were conducted to assess the significance of differences in IC50 values between single-agent (Cis or 5-FU) and combination treatments. A highly significant difference (****P < 0.0001) was observed, indicating a strong level of statistical confidence in the observed effects.
The normalized isobologram in Figure S4 (Supplementary file 1) illustrates the interaction of bergenin combined with Cis and 5-FU at non-constant ratios. This analysis maps out various points, representing different combination groups, which exhibit antagonistic, additive, or synergistic effects. Isobolograms are typically used to visualize drug interactions, where points below the line indicate synergy, points on the line suggest additivity, and those above the line indicate antagonism. This isobologram reveals that most of the combination groups fall below the line, showing a predominantly synergistic interaction of bergenin in combination with Cis and 5-FU in both MCF-7 and HCT 116 cell lines, respectively. Combination groups as shown in Table 4, were finalized based on the lowest Combination index at 50% inhibition effect. Based on the CI value, it can be inferred that both groups show synergistic behavior in reducing cell viability and proliferation.
Table 4.
Doses (μM) of combination groups and combination indices at ≈50% effect
|
Combination groups
|
Combination doses
|
Fa (≈ 50 % effect)
|
Combination index (CI)
|
Description based on CI
|
| Bergenin + Cis |
20 μM + 5 μM |
0.537 |
0.897 |
Synergism |
| Bergenin + 5-FU |
10 μM + 10 μM |
0.495 |
0.684 |
Synergism |
Fluorescent staining
Dual AO/EtBr (acridine orange/ethidium bromide) staining
Acridine orange has the ability to permeate normal cell membranes. In the absence of any treatment, cells exhibit a bright green fluorescence, highlighting a well-organized structure. Conversely, early apoptotic cells display a yellow-orange hue, while late apoptotic cells, marked by nuclear shrinkage, damage, and blebbing, exhibit an orange-red color. Cells with red-colored nuclei at higher concentrations suggest late-stage apoptosis. Untreated cells, maintain their healthy appearance with a vibrant green fluorescence. Compared to the untreated control, the 24 hour treated groups (bergenin and Cis alone as well as in combination) exhibited distinctive apoptotic features such as blebbing, chromatin fragmentation, condensed chromatin, and nuclear shrinkage (Figure 2). Untreated HCT 116 cells when compared with treated groups (bergenin and 5-FU alone as well as in combination), also showed similar alterations after 24 hours treatment as highlighted in Figure 3.
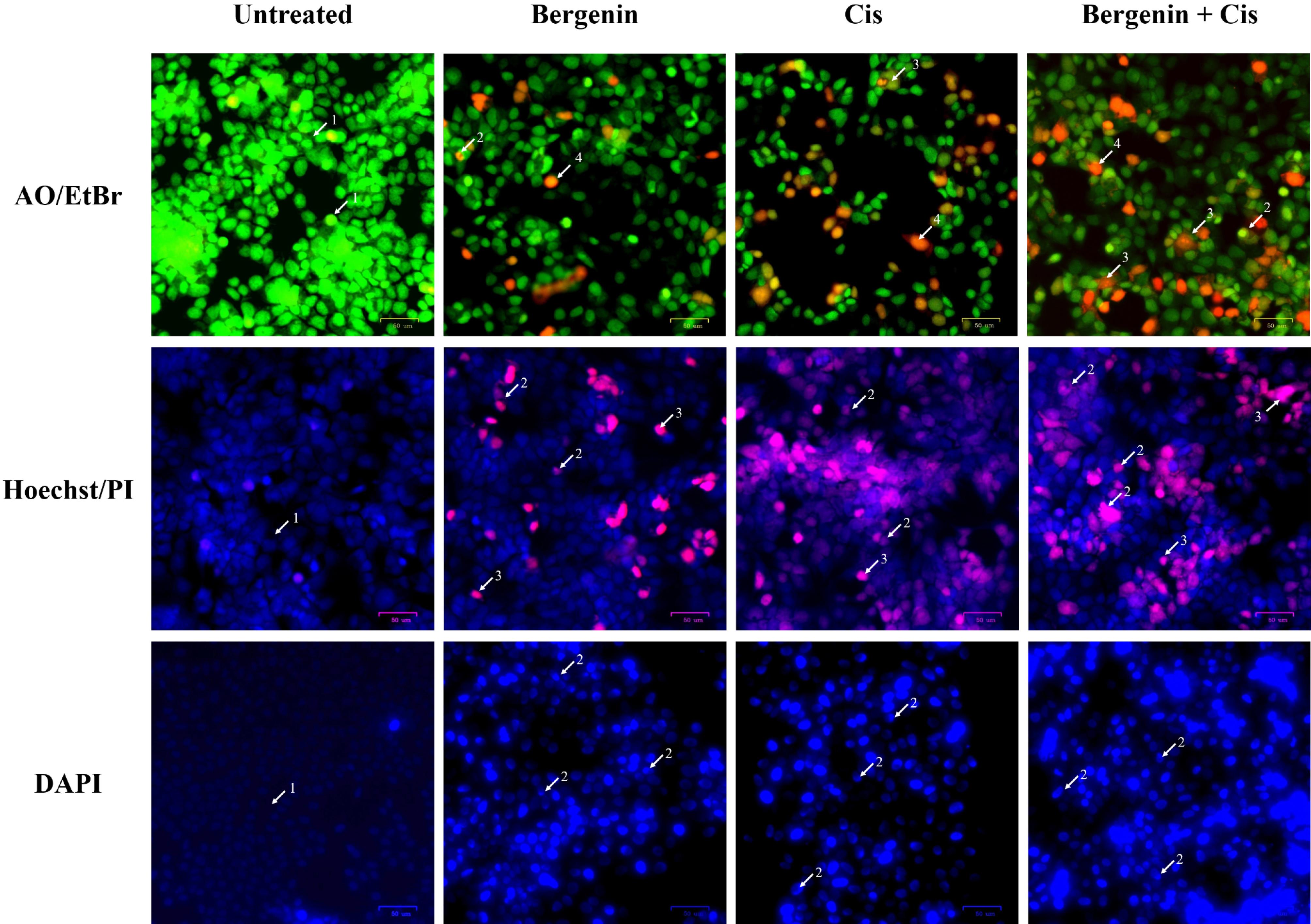
Figure 2.
Fluorescent staining micrographs of treated MCF-7 cells compared to untreated cells. Morphological changes are highlighted using white arrows for AO/EtBr staining (1. Viable cells with bright green nuclei, 2. Early apoptotic cells with fragmented green nuclei, 3. Late apoptotic cells with red fragmented nuclei, 4. Dead cells, 5. Apoptotic bodies); Hoechst/PI staining (1. Viable cells with intact blue nuclei, 2. Apoptotic cells with pink fragmented nuclei and apoptotic bodies, 3. Dead cells); and DAPI staining (1. Viable cells with intact nuclei, 2. Apoptotic cells with fragmented bright blue nuclei)
.
Fluorescent staining micrographs of treated MCF-7 cells compared to untreated cells. Morphological changes are highlighted using white arrows for AO/EtBr staining (1. Viable cells with bright green nuclei, 2. Early apoptotic cells with fragmented green nuclei, 3. Late apoptotic cells with red fragmented nuclei, 4. Dead cells, 5. Apoptotic bodies); Hoechst/PI staining (1. Viable cells with intact blue nuclei, 2. Apoptotic cells with pink fragmented nuclei and apoptotic bodies, 3. Dead cells); and DAPI staining (1. Viable cells with intact nuclei, 2. Apoptotic cells with fragmented bright blue nuclei)
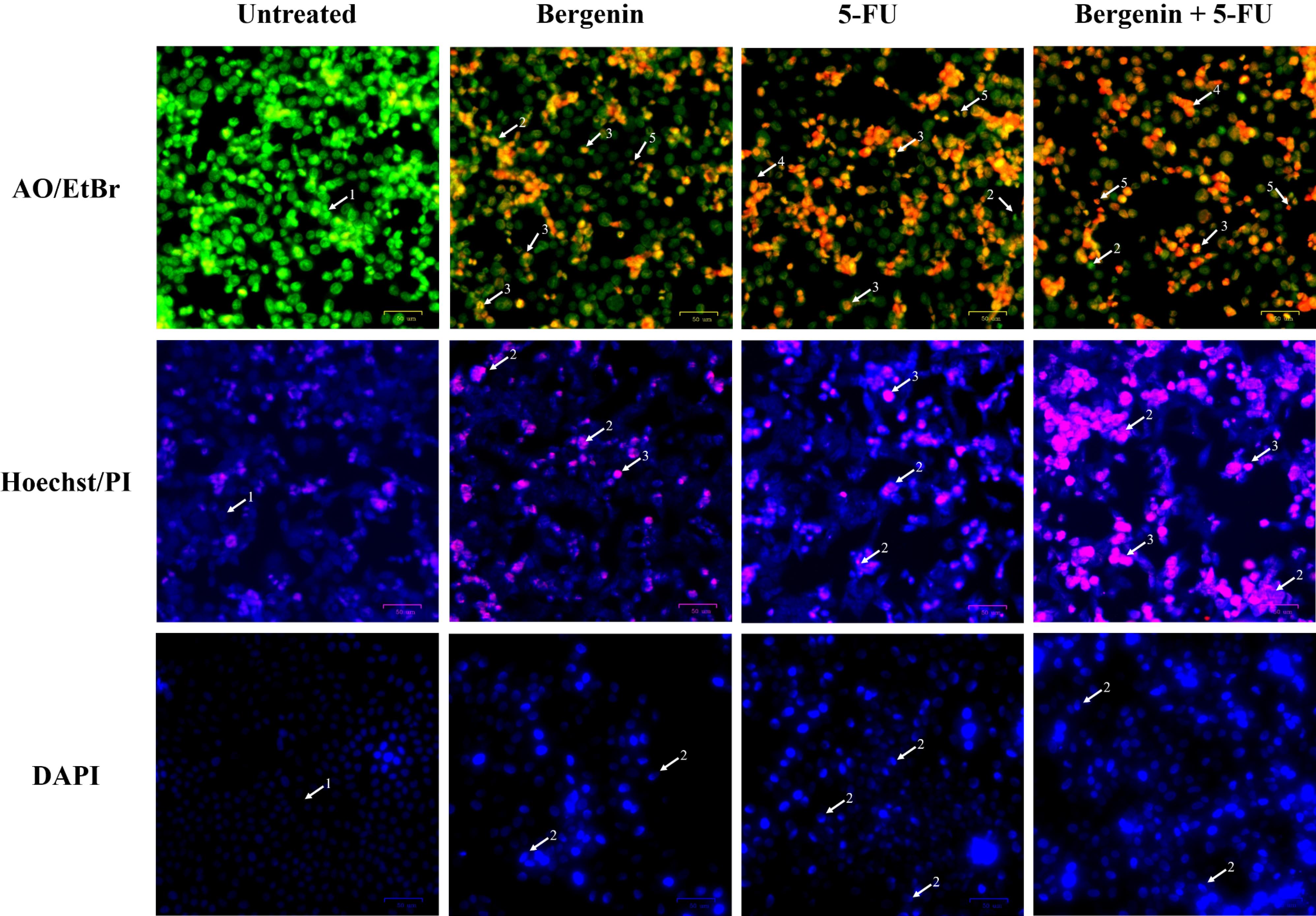
Figure 3.
Fluorescent staining micrographs of treated HCT 116 cells compared to untreated cells. Morphological changes are highlighted using white arrows for AO/EtBr staining (1. Viable cells with bright green nuclei, 2. Early apoptotic cells with fragmented green nuclei, 3. Late apoptotic cells with red fragmented nuclei, 4. Dead cell, 5. Apoptotic bodies); Hoechst/PI staining (1. Viable cells with intact blue nuclei, 2. Apoptotic cells with pink fragmented nuclei, 3. Dead cells); and DAPI staining (1. Viable cells with intact nuclei, 2. Apoptotic cells with fragmented bright blue nuclei)
.
Fluorescent staining micrographs of treated HCT 116 cells compared to untreated cells. Morphological changes are highlighted using white arrows for AO/EtBr staining (1. Viable cells with bright green nuclei, 2. Early apoptotic cells with fragmented green nuclei, 3. Late apoptotic cells with red fragmented nuclei, 4. Dead cell, 5. Apoptotic bodies); Hoechst/PI staining (1. Viable cells with intact blue nuclei, 2. Apoptotic cells with pink fragmented nuclei, 3. Dead cells); and DAPI staining (1. Viable cells with intact nuclei, 2. Apoptotic cells with fragmented bright blue nuclei)
Dual propidium iodide/Hoechst 33342 staining
The Hoechst 33342 dye, which emits blue fluorescence, serves as a cell-permeable nucleic acid stain commonly used for identifying chromatin condensation and fragmentation. It specifically stains the fragmented nuclei of cells undergoing apoptosis. Conversely, the pink fluorescent PI functions as a DNA-binding dye that selectively enters cells with compromised plasma membrane integrity, indicating instances of increased permeability. Treatment-induced apoptosis in MCF-7 and HCT 116 cells were characterized by morphological features of apoptosis, including chromatin condensation and nuclear fragmentation (marked by arrows in Figures 2 and 3).
DAPI (4′,6-diamidine-2′-phenylindole dihydrochloride) staining
Untreated MCF-7 and HCT 116 cells showed faint, homogenous blue fluorescent nuclei, suggesting intact nuclear structure. The signs of apoptosis were then observed when MCF-7 cells treated bergenin and Cis alone as well as in combination for 24 hours, the blue emission was much brighter than in the unaffected cells, indicating features of apoptosis like chromatin fragmentation and condensation (Figure 2). Moreover, as shown in Figure 3, the treated HCT 116 cells (bergenin and 5-FU alone and their combination) displayed nuclear fragmentation, confirming the induction of apoptosis compared to untreated cells.
Colony forming assay
Treatment with bergenin, both individually and in conjunction with Cis and 5-FU, resulted in significant inhibitory effects on colony formation. Figure 4 depicts a notable reduction in colony formation of MCF-7 and HCT 116 cancer cells following 24 hours of treatment with bergenin alone and in combination with Cis and 5-FU, respectively. In MCF-7 cells, the combined treatment of bergenin and Cis exhibited a substantial decrease in colony formation (53.57%; P < 0.0001) compared to untreated groups. Similarly, bergenin and Cis monotherapies independently prevented colony formation, 67% (P < 0.0001) and 64% (P < 0.0001) area of colonies observed, respectively, when compared to the untreated group. In HCT 116 cells, as shown in Figure 4, combined treatment with bergenin and 5-FU significantly inhibited colony formation, reducing the number of colonies to 33.46% (P < 0.0001) compared to the untreated group. Treatment with bergenin alone resulted in a significant reduction to 83.58% (P < 0.0001), while treatment with 5-FU alone reduced the colony count to 55.93% (P < 0.0001), both compared to the untreated group.
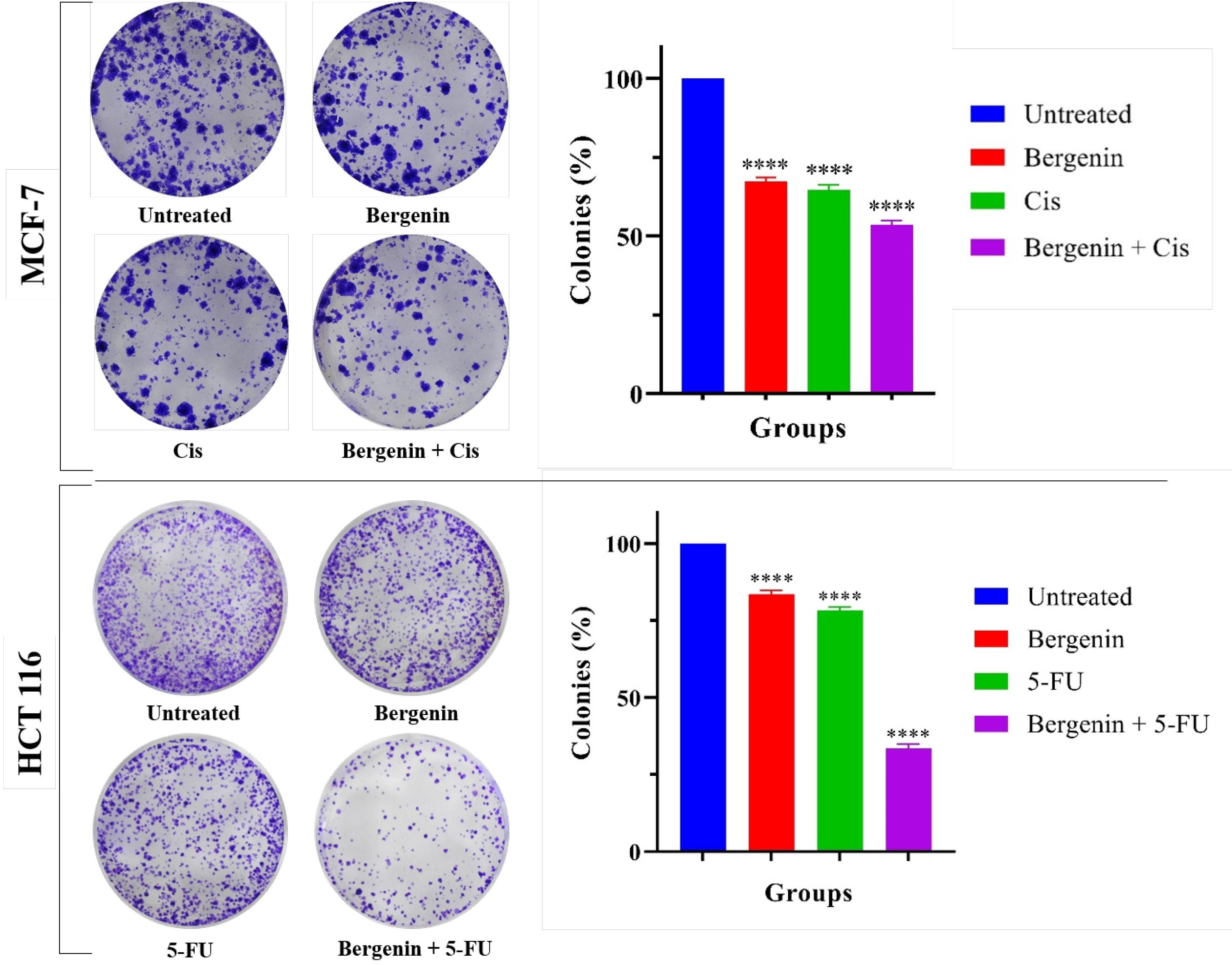
Figure 4.
Colony forming assay on MCF-7 and HCT 116 cells, significant reduction in colony number was observed in treated groups when compared to untreated; Images of colonies and bar graph showing the number of colonies as survival fraction compared to untreated groups. All experiments were performed independently in triplicate. Values are expressed in mean ± SD. Statistical analysis identified a highly significant difference between the treated and untreated groups (****P < 0.0001), demonstrating a strong level of confidence that the observed effect is unlikely to be attributable to chance
.
Colony forming assay on MCF-7 and HCT 116 cells, significant reduction in colony number was observed in treated groups when compared to untreated; Images of colonies and bar graph showing the number of colonies as survival fraction compared to untreated groups. All experiments were performed independently in triplicate. Values are expressed in mean ± SD. Statistical analysis identified a highly significant difference between the treated and untreated groups (****P < 0.0001), demonstrating a strong level of confidence that the observed effect is unlikely to be attributable to chance
Wound healing assay
The results revealed a noteworthy reduction in the cell migration capacity of MCF-7 and HCT 116 cells following a 72 hours treatment, in comparison to the untreated group. Figure 5 displays images of wound healing assay for MCF-7 and HCT 116 cells at 0, 24, 48, and 72 hours for group I to VIII.
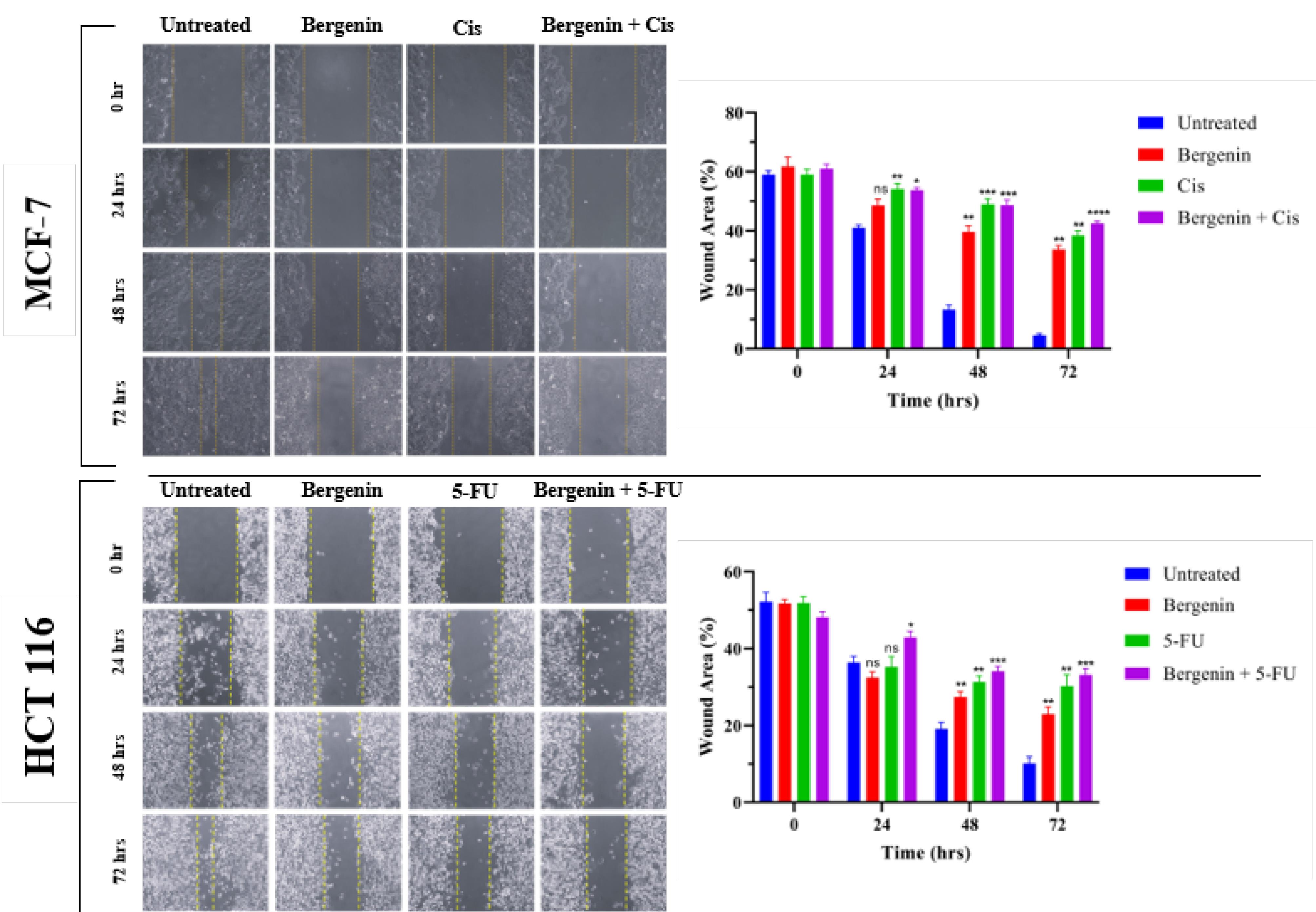
Figure 5.
The wound healing assay conducted on MCF-7 and HCT 116 cells included - capturing images of wound closure at 24-hrs intervals over a 72-hrs period, and bar graphs depicting the relationship between wound area (%) and time. The results indicated a significant decrease in cell migration for treated groups when compared to untreated groups. All results were averaged for triple independent experiments. Values are expressed in mean ± SD. Statistical significance was considered as *P < 0.05; **P < 0.01; ***P < 0.001; and ****P < 0.0001 when all treated groups compared to untreated group
.
The wound healing assay conducted on MCF-7 and HCT 116 cells included - capturing images of wound closure at 24-hrs intervals over a 72-hrs period, and bar graphs depicting the relationship between wound area (%) and time. The results indicated a significant decrease in cell migration for treated groups when compared to untreated groups. All results were averaged for triple independent experiments. Values are expressed in mean ± SD. Statistical significance was considered as *P < 0.05; **P < 0.01; ***P < 0.001; and ****P < 0.0001 when all treated groups compared to untreated group
MCF-7
In the untreated group, the wound completely closed within 72 hours as shown in Figure 5, with the wound area decreasing from 53.72% (0 hours) to 4.949% (72 hours). In contrast, the groups treated with Cis alone, bergenin alone, or the combination of bergenin and Cis showed only a minimal reduction in wound area. Statistical analysis revealed a significant inhibition in cell migration for all three treatment groups. The wound area in cells treated with bergenin alone decreased to 39.65% (P < 0.001) and to 48.94% (P < 0.001) when combined with Cis after 48 hours, compared to the untreated group. After 72 hours, cells treated with bergenin alone showed a wound area of 38.44% (P < 0.01), while those treated with the bergenin-Cis combination had a wound area of 42.51% (P < 0.0001), showing significant inhibition of cell migration compared to the control group.
HCT 116
As shown in Figure 5, the untreated HCT 116 cells exhibited a substantial decrease in width of the wound, declining from 52.27% (0 hours) to 19.14% after 48 hours and further to 10.13% after 72 hours. The positive control, 5-FU impeded wound closure, resulting in a reduction of gap size only from 51.86% at 0 hours to 29.94% after 48 hours and to 28.58% after 72 hours. Notably, when compared to both bergenin alone and its combination with 5-FU, there was a significant reduction in wound gap to 27.43% (P < 0.001) and 34.17% (P < 0.0001) after 48 hours, and further to 22.91% (P < 0.01) and 33.17% (P < 0.0001) after 72 hours, respectively.
Cell cycle analysis
Flow cytometry was utilized to analyze the cell cycle of MCF-7 and HCT 116 cells, aiming to ascertain whether cell cycle arrest contributed to the reduction in cell density.
MCF-7
As depicted in Figure 6, after 24 hours the cell cycle retardation was observed during the culture period, in both the treated and untreated cells of MCF-7. Results showed that 72.43%, 6.61% and 20.96% of untreated cells were in G0/G1, S and G2/M phase, respectively. Treatment of bergenin and Cis alone significantly retarded the cells at G0/G1 phase with 80.66% (P < 0.01) and 84.19% (P < 0.001) cells respectively compared to untreated. When exposed to bergenin, 6.53% and 12.81% of cells were found to be confined in the S and G2/M phases, respectively. Conversely, treatment with Cis resulted in 7.36% and 8.45% of cells categorized in the S and G2/M phases, respectively. While treatment of bergenin and Cis in combination significantly retarded 83.10% (P < 0.001) cells in G0/G1, 7.75% in S and 9.15% in G2/M phase.
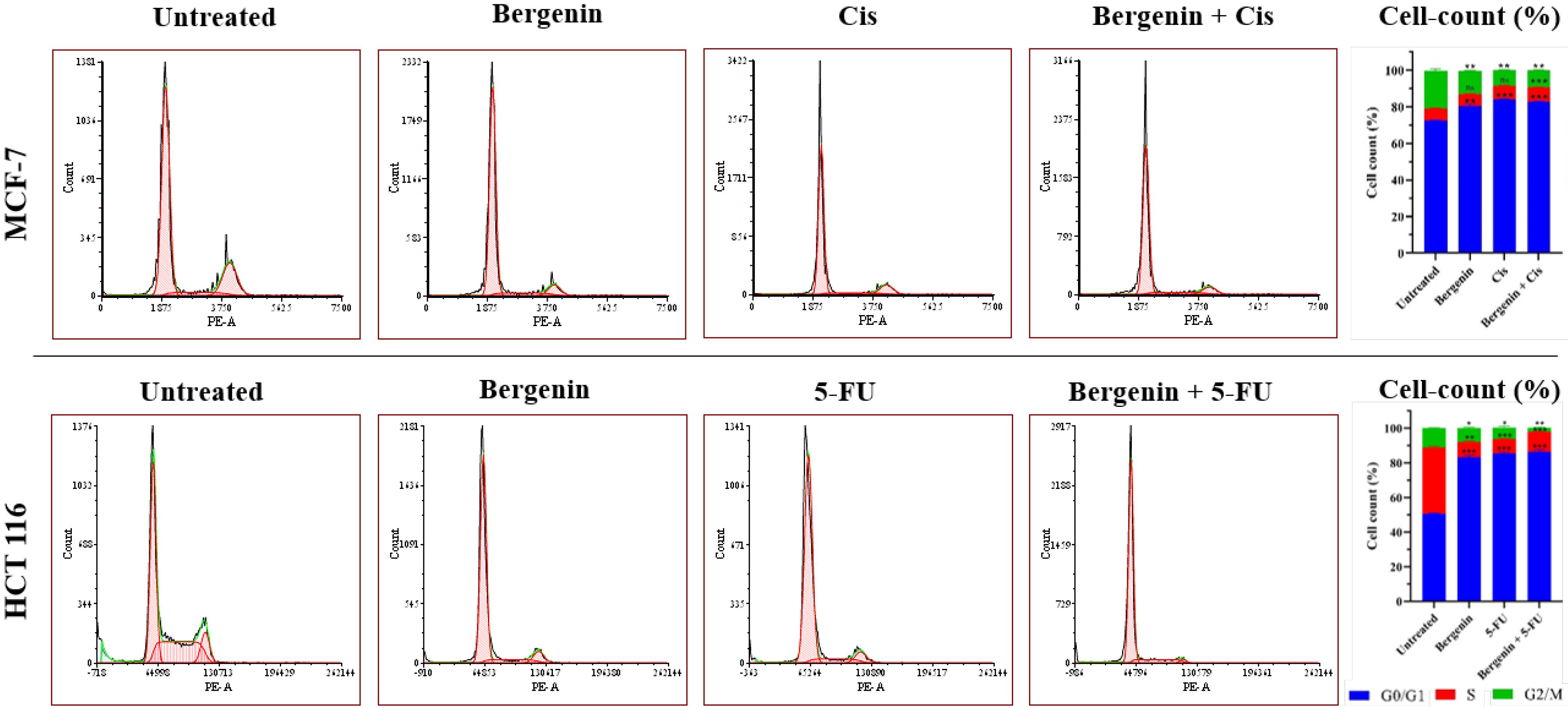
Figure 6.
Results of cell cycle analysis for groups I to IV and groups V to VIII on MCF-7 and HCT 116 cells respectively, represented as cell count plotted against DNA content (PI fluorescence/cell); and percent cell count, where cell count (%) at G2/M stage is significantly low in treated groups compared to the untreated groups (*P < 0.05; **P < 0.01 and ***P < 0.001). Values are expressed in mean ± SD
.
Results of cell cycle analysis for groups I to IV and groups V to VIII on MCF-7 and HCT 116 cells respectively, represented as cell count plotted against DNA content (PI fluorescence/cell); and percent cell count, where cell count (%) at G2/M stage is significantly low in treated groups compared to the untreated groups (*P < 0.05; **P < 0.01 and ***P < 0.001). Values are expressed in mean ± SD
HCT 116
As depicted in Figure 6, both treated and untreated HCT 116 cells exhibited a delay in cell cycle progression after 24 hours. The findings revealed that, 50.55% of the untreated cells were in G0/G1 phase, while 38.19% were in S phase, and 11.27% were in G2/M phase. Separate treatment of bergenin and 5-FU led to significant cell cycle arrest with 83.14% (P < 0.001) and 85.18% (P < 0.001) of cells primarily in the G0/G1 phase, respectively. Exposure to bergenin resulted in 8.77% of cells in the S phase and 8.09% in the G2/M phase while 5-FU treatment led to 8.32% of cells in the S phase and 6.50% in the G2/M phase. The combination dose of bergenin and 5-FU significantly arrested 86.44% (P < 0.001) cells at G0/G1 phase, and only a few cells (11.54% and 2.02%) were in S and G2/M phase, respectively. These findings suggest that bergenin may inhibit cell cycle progression of both MCF-7 and HCT 116 cells, primarily by arresting cells at the G0/G1 phase.
Cell apoptosis analysis
MCF-7
It was observed (Figure 7) in flow cytometry analysis for apoptosis that 87.66% of cells in the untreated MCF-7 cell group were live, with only 12.33% identified as early apoptotic cells. Very few cells were detected in the other two quadrants, which represent dead and late apoptotic cells. Upon treatment with the positive control Cis, there was a significant shift in cell distribution towards the early apoptotic quadrant, with 75.42% (P < 0.0001) of cells falling into this category, while the percentage of live cells decreased to 18.93% (P < 0.0001) as compared to untreated control group. Similar induction of apoptosis was observed in the group treated with bergenin alone, with 67.21% (P < 0.0001) of cells in early apoptosis and 9.72% (P < 0.0001) in late apoptosis, while the remaining 23.08% (P < 0.0001) were live as compared to untreated control group. Furthermore, when cells were treated with both bergenin and Cis in combination, there was a significant increase in early apoptotic cells, accounting for 81.89% (P < 0.0001) of the total, while 7.78% (P < 0.0001) were in the late apoptotic stage when compared to untreated control group.
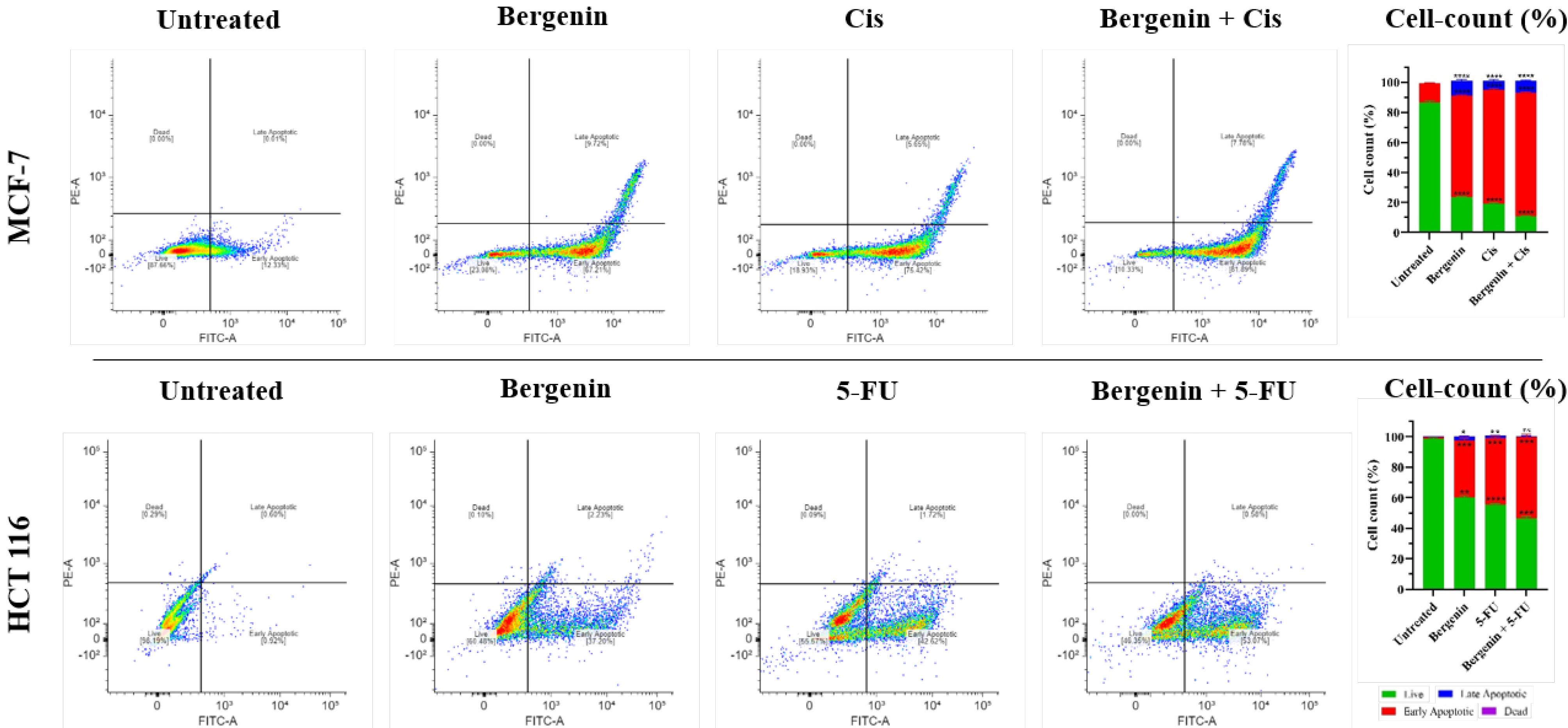
Figure 7.
Apoptosis detection assay on MCF-7 and HCT 116 cells revealed that percent of early apoptotic cells increased when treated with Cis or 5-FU and Bergenin alone and in combination respectively; increased percent of early apoptotic cells as shown in bar graphs was statistically significant (*P < 0.05; **P < 0.01; ***P < 0.001 and ****P < 0.0001) in all treatment groups when compared to untreated groups. Values are expressed in mean ± SD
.
Apoptosis detection assay on MCF-7 and HCT 116 cells revealed that percent of early apoptotic cells increased when treated with Cis or 5-FU and Bergenin alone and in combination respectively; increased percent of early apoptotic cells as shown in bar graphs was statistically significant (*P < 0.05; **P < 0.01; ***P < 0.001 and ****P < 0.0001) in all treatment groups when compared to untreated groups. Values are expressed in mean ± SD
HCT 116
Flow cytometric analysis, as shown in Figure 7, demonstrated that 99.17% of cells in untreated HCT 116 cells were viable, with only 0.42%, 0.04%, and 0.38% in early apoptotic, late apoptotic, and dead states, respectively.Upon treatment with 5-FU and bergenin individually, there was a significant increase in early apoptotic cells to 40.44% (P < 0.0001) and 23.87% (P < 0.0001), respectively, compared to the untreated group. Similarly, the group treated with both bergenin, and 5-FU exhibited a significant increase in apoptotic cells to 40.87% (P < 0.0001) compared to the untreated group. This data suggests that bergenin alone and in combination with Cis or 5-FU has the potential to induce apoptosis in MCF-7 and HCT 116 cells, respectively.
Discussion
Bergenin can be useful to reduce the cancer cell viability.21,22 The present study on breast and colorectal cancer cells (MCF-7 and HCT 116 respectively) also demonstrated the concentration and time-dependent significant reduction in cell viability. Combination analysis established initial dosage parameters for its synergistic use with Cis and 5-FU. CompuSyn analysis12 confirmed that bergenin co-administration reduced the required doses of these chemotherapeutic agents while enhancing their efficacy, achieving optimal inhibition at lower concentrations.
For MCF-7 cells, a 50% viability (IC50 value) reduction required 34.16 µM of bergenin or 14.23 µM of Cis alone, while in best reduced combination dose, only 20 µM of bergenin and 5 µM of Cis were needed. Similarly, in HCT 116 cells, 40.93 µM of bergenin or 22.87 µM of 5-FU alone achieved a 50% reduction, while the best reduced combination dose required just 10 µM of each. This synergy reduces the need for higher chemotherapy doses, potentially mitigating side effects. Similar synergistic effects were observed in other studies combining Cis with piperine23 and curcumin24 in MCF-7 cells and 5-FU with diosmetin25, apigenin26, and zerumbone27 in HCT 116 cells.
Various in vitro tests were conducted to evaluate the inhibitory effects of bergenin, Cis, 5-FU, and their combinations on MCF-7 and HCT 116 cells. The analysis revealed that bergenin, both alone and in combination with 5-FU or Cis, effectively halted cell cycle progression at the G0/G1 phase. This interruption is critical as it prevents cancer cells from synthesizing DNA and progressing to mitosis, thereby inhibiting their proliferation. This effect is likely due to the downregulation of cyclins and cyclin-dependent kinases needed for the G1 to S phase transition,28 underscoring bergenin’s potential to disrupt cell cycle dynamics in cancer cells.
The colony formation and wound healing assays further showed the reduction in cells’ capacity for unlimited proliferation and migration potential post-treatment. Notably, treatment with bergenin, both alone and in combination with Cis or 5-FU, significantly suppressed the clonogenic formation of MCF-7 and HCT 116 cells, respectively. The findings of this study also supported previous experiments that showed reduced colony formation and inhibited wound closure in human cancer cells treated with various combinations.29-33 These inhibitory effects are likely mediated through the induction of apoptosis, which is important for eliminating cancer cells. Apoptosis acts as a crucial defense mechanism against carcinogenesis, eliminating genetically defective cells.34,35 Therefore, inducing apoptosis is highly desirable in cancer management.36-38 The present study not only proves the apoptosis of cell using fluorescent staining (AO/EtBr, DAPI, and Hoechst/PI staining) but also proves it using flow cytometry study. AO/EtBr staining differentiated between live, early apoptotic, and late apoptotic cells, while DAPI was used to observe nuclear integrity and fragmentation in the treatment groups. Hoechst/PI dual staining further differentiated nuclear damage by identifying condensed pyknotic nuclei with Hoechst and distinguishing late apoptotic from normal cells using PI in treated cells. To further quantify the apoptotic potential of bergenin and the combination treatments, the expression of annexin V, a protein that binds tightly to PS in a calcium-dependent manner, was evaluated using flow cytometry. These fluorescent staining and flow cytometry analysis prove that bergenin, alone and in combination with 5-FU or Cis, notably increased the proportion of apoptotic cells in both MCF-7 and HCT 116. Similar study carried out using bergenin and other phytocomponents also support these findings and prove that apoptosis is the major activity to eliminate the compromised cells.39-41
Cancer drugs induce cell death through apoptosis, with their efficacy directly related to the level of apoptosis induced.42,43 This pro-apoptotic effect may involve intrinsic pathways characterized by mitochondrial dysfunction and caspase activation, as well as extrinsic pathways involving death receptors.44-46 Recent studies emphasize combining natural compounds with conventional chemotherapy to enhance efficacy and reduce toxicity. Gano and colleagues46 demonstrated synergistic phytochemical combinations in prostate cancer, while Hakeem et al47 reported piperine’s ability to enhance Doxorubicin’s effects in breast cancer. Detailed in vivo and in vitro phytochemical studies against breast and colorectal cancers carried out by Talib and co-workers48 also corroborate the findings of present study. Clinical investigations on tamoxifen-thymoquinone49 and on sulforaphane-docetaxel,50 further support the role of phytochemicals in chemotherapy enhancement. More recently, Li et al51 demonstrated that ginsenosides synergize with Cis, improving efficacy and reducing toxicity. These findings align with broader research on Cis-based combination therapies, which enhance anticancer effects while mitigating drug resistance and adverse effects. Collectively, these insights support bergenin as a promising candidate for combination therapy, reducing chemotherapy-associated toxicity while maintaining efficacy and helping to improve patient outcomes and quality of life. By effectively reducing the required dosages of traditionally applied chemotherapeutic agents without compromising their efficacy, bergenin could potentially be helpful to reduce the adverse side effects associated with high-dose chemotherapy.
Conclusion
These findings show that bergenin plays a dual role, inhibiting the progression of cancer cells while promoting apoptosis. Furthermore, when combined with 5-FU or Cis, it synergistically inhibits cancer cell proliferation and enhances the apoptotic potential of Cis and 5-FU, even at lower doses. This combination approach alleviates the adverse effects linked to higher doses of Cis and 5-FU in the treatment of breast and colorectal cancer. Therefore, utilizing bergenin alongside chemotherapy drugs can minimize their dose and side effects and improve treatment outcomes through synergistic effects.
Competing Interests
The authors declare that they have no conflict of interest.
Supplementary Files
Supplementary file 1 contains Figures S1-S4.
(pdf)
Acknowledgements
The first author expresses sincere gratitude to the Council of Scientific & Industrial Research (CSIR), Ministry of Science and Technology, Government of India, for support of the author’s doctoral research (Ph.D.) [Award No. 09/0070(11301)/2021-EMR-I].
References
- Ferlay J, Ervik M, Lam F, Laversanne M, Colombet M, Mery L, et al. Global Cancer Observatory: Cancer Today. Lyon, France: International Agency for Research on Cancer; 2024. Available from: https://gco.iarc.who.int/today. Accessed January 18, 2025.
- Sultana T, Mitra AK, Das S. Evaluation of anti-cancer potential of Excoecariaagallocha (L) leaf extract on human cervical cancer (SiHa) cell line and assessing the underlying mechanism of action. Futur J Pharm Sci 2022; 8(1):3. doi: 10.1186/s43094-021-00389-y [Crossref] [ Google Scholar]
- Chauhan R, Malhotra L, Gupta A, Dagar G, Mendiratta M, Masoodi T. Bergenin inhibits growth of human cervical cancer cells by decreasing galectin-3 and MMP-9 expression. Sci Rep 2024; 14(1):15287. doi: 10.1038/s41598-024-64781-3 [Crossref] [ Google Scholar]
- Gan Y, Li X, Han S, Zhou L, Li W. Targeting Mcl-1 degradation by bergenin inhibits tumorigenesis of colorectal cancer cells. Pharmaceuticals (Basel) 2023; 16(2):241. doi: 10.3390/ph16020241 [Crossref] [ Google Scholar]
- Li X, Xie L, Zhou L, Gan Y, Han S, Zhou Y. Bergenin inhibits tumor growth and overcomes radioresistance by targeting aerobic glycolysis. Am J Chin Med 2023; 51(7):1905-25. doi: 10.1142/s0192415x23500842 [Crossref] [ Google Scholar]
- Li X, Gan Y, Han S, Liao J, Zhou L, Li W. Bergenin Exerts Antitumor Effects by Promoting Survivin Degradation. Res Sq [Preprint]. April 14, 2022 [Cited 2024 July 23]. Available from: https://www.researchsquare.com/article/rs-1547129/v1.
- Liu Y, An Z, He Y. The traditional uses, phytochemistry, pharmacology and toxicology of Bergenia purparescens: a review comments and suggestions. Heliyon 2023; 9(11):e22249. doi: 10.1016/j.heliyon.2023.e22249 [Crossref] [ Google Scholar]
- Dasari S, Tchounwou PB. Cisplatin in cancer therapy: molecular mechanisms of action. Eur J Pharmacol 2014; 740:364-78. doi: 10.1016/j.ejphar.2014.07.025 [Crossref] [ Google Scholar]
- Lembersky BC, Wieand HS, Petrelli NJ, O’Connell MJ, Colangelo LH, Smith RE. Oral uracil and tegafur plus leucovorin compared with intravenous fluorouracil and leucovorin in stage II and III carcinoma of the colon: results from National Surgical Adjuvant Breast and Bowel Project Protocol C-06. J Clin Oncol 2006; 24(13):2059-64. doi: 10.1200/jco.2005.04.7498 [Crossref] [ Google Scholar]
- Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 1983; 65(1-2):55-63. doi: 10.1016/0022-1759(83)90303-4 [Crossref] [ Google Scholar]
- Chou TC. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol Rev 2006; 58(3):621-81. doi: 10.1124/pr.58.3.10 [Crossref] [ Google Scholar]
- Chou TC, Talalay P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv Enzyme Regul 1984; 22:27-55. doi: 10.1016/0065-2571(84)90007-4 [Crossref] [ Google Scholar]
- Gherghi I, Girousi ST, Voulgaropoulos AN, Tzimou-Tsitouridou R. Study of interactions between DNA-ethidium bromide (EB) and DNA-acridine orange (AO), in solution, using hanging mercury drop electrode (HMDE). Talanta 2003; 61(2):103-12. doi: 10.1016/s0039-9140(03)00238-8 [Crossref] [ Google Scholar]
- Baskić D, Popović S, Ristić P, Arsenijević NN. Analysis of cycloheximide-induced apoptosis in human leukocytes: fluorescence microscopy using annexin V/propidium iodide versus acridin orange/ethidium bromide. Cell Biol Int 2006; 30(11):924-32. doi: 10.1016/j.cellbi.2006.06.016 [Crossref] [ Google Scholar]
- Cao H, Li C, Qi W, Meng X, Tian R, Qi Y. Synthesis, cytotoxicity and antitumour mechanism investigations of polyoxometalate doped silica nanospheres on breast cancer MCF-7 cells. PLoS One 2017; 12(7):e0181018. doi: 10.1371/journal.pone.0181018 [Crossref] [ Google Scholar]
- Atale N, Chakraborty M, Mohanty S, Bhattacharya S, Nigam D, Sharma M. Cardioprotective role of Syzygiumcumini against glucose-induced oxidative stress in H9C2 cardiac myocytes. Cardiovasc Toxicol 2013; 13(3):278-89. doi: 10.1007/s12012-013-9207-1 [Crossref] [ Google Scholar]
- Kabakov AE, Kudryavtsev VA, Gabai VL. Determination of cell survival or death. Methods Mol Biol 2011; 787:231-44. doi: 10.1007/978-1-61779-295-3_17 [Crossref] [ Google Scholar]
- Jonkman JE, Cathcart JA, Xu F, Bartolini ME, Amon JE, Stevens KM. An introduction to the wound healing assay using live-cell microscopy. Cell Adh Migr 2014; 8(5):440-51. doi: 10.4161/cam.36224 [Crossref] [ Google Scholar]
- Chung SS, Dutta P, Austin D, Wang P, Awad A, Vadgama JV. Combination of resveratrol and 5-flurouracil enhanced anti-telomerase activity and apoptosis by inhibiting STAT3 and Akt signaling pathways in human colorectal cancer cells. Oncotarget 2018; 9(68):32943-57. doi: 10.18632/oncotarget.25993 [Crossref] [ Google Scholar]
- Jin S, Zhang QY, Kang XM, Wang JX, Zhao WH. Daidzein induces MCF-7 breast cancer cell apoptosis via the mitochondrial pathway. Ann Oncol 2010; 21(2):263-8. doi: 10.1093/annonc/mdp499 [Crossref] [ Google Scholar]
- Mehta S, Kadian V, Dalal S, Dalal P, Kumar S, Garg M. A fresh look on bergenin: vision of its novel drug delivery systems and pharmacological activities. Future Pharmacol 2022; 2(1):64-91. doi: 10.3390/futurepharmacol2010006 [Crossref] [ Google Scholar]
- Kumar P, Ansari K, Jhala D. Deciphering the therapeutic potential of bergenin in breast cancer: in silico insights into HSP90AA1 and HRAS interaction mediated inhibition of PI3K-Akt and MAPK signaling pathway. Indian J Biochem Biophys 2024; 61(11):639-58. doi: 10.56042/ijbb.v61i11.6391 [Crossref] [ Google Scholar]
- Fattah A, Morovati A, Niknam Z, Mashouri L, Asadi A, Tvangar Rizi S. The synergistic combination of cisplatin and piperine induces apoptosis in MCF-7 cell line. Iran J Public Health 2021; 50(5):1037-47. doi: 10.18502/ijph.v50i5.6121 [Crossref] [ Google Scholar]
- Zou J, Zhu L, Jiang X, Wang Y, Wang Y, Wang X. Curcumin increases breast cancer cell sensitivity to cisplatin by decreasing FEN1 expression. Oncotarget 2018; 9(13):11268-78. doi: 10.18632/oncotarget.24109 [Crossref] [ Google Scholar]
- Kamran S, Sinniah A, Chik Z, Alshawsh MA. Diosmetin exerts synergistic effects in combination with 5-fluorouracil in colorectal cancer cells. Biomedicines 2022; 10(3):531. doi: 10.3390/biomedicines10030531 [Crossref] [ Google Scholar]
- Yang C, Song J, Hwang S, Choi J, Song G, Lim W. Apigenin enhances apoptosis induction by 5-fluorouracil through regulation of thymidylate synthase in colorectal cancer cells. Redox Biol 2021; 47:102144. doi: 10.1016/j.redox.2021.102144 [Crossref] [ Google Scholar]
- Dehghan R, Bahreini F, Najafi R, Saidijam M, Amini R. The combination of zerumbone and 5‐FU: a significant therapeutic strategy in sensitizing colorectal cancer cells to treatment. Biomed Res Int 2021; 2021(1):6635874. doi: 10.1155/2021/6635874 [Crossref] [ Google Scholar]
- Ding L, Cao J, Lin W, Chen H, Xiong X, Ao H. The roles of cyclin-dependent kinases in cell-cycle progression and therapeutic strategies in human breast cancer. Int J Mol Sci 2020; 21(6):1960. doi: 10.3390/ijms21061960 [Crossref] [ Google Scholar]
- Kwon H, Kim Y, Kim JH. A combination of myokines and genistein suppresses cancer stemness in MCF-7 human breast cancer cells. Nutr Res Pract 2024; 18(3):436-45. doi: 10.4162/nrp.2024.18.3.436 [Crossref] [ Google Scholar]
- El-Far AH, Saddiq AA, Mohamed SA, Almaghrabi OA, Mousa SA. Curcumin and thymoquinone combination attenuates breast cancer cell lines’ progression. Integr Cancer Ther 2022; 21:15347354221099537. doi: 10.1177/15347354221099537 [Crossref] [ Google Scholar]
- Ghosh S, Pal A, Ray M. 5FU synergistically inhibits MCF-7 in combination with methylglyoxal. Clin Oncol 2017; 2:1353. [ Google Scholar]
- Nurhayati IP, Khumaira A, Ilmawati GPN, Meiyanto E, Hermawan A. Cytotoxic and antimetastatic activity of hesperetin and doxorubicin combination toward Her2 expressing breast cancer cells. Asian Pac J Cancer Prev 2020; 21(5):1259-67. doi: 10.31557/apjcp.2020.21.5.1259 [Crossref] [ Google Scholar]
- Kanno SI, Yomogida S, Tomizawa A, Yamazaki H, Ukai K, Mangindaan RE. Combined effect of papuamine and doxorubicin in human breast cancer MCF-7 cells. Oncol Lett 2014; 8(2):547-50. doi: 10.3892/ol.2014.2218 [Crossref] [ Google Scholar]
- Morana O, Wood W, Gregory CD. The apoptosis paradox in cancer. Int J Mol Sci 2022; 23(3):1328. doi: 10.3390/ijms23031328 [Crossref] [ Google Scholar]
- Thapa S, Rather RA, Singh SK, Bhagat M. Insights into the role of defective apoptosis in cancer pathogenesis and therapy. In: Tutar Y, ed. Regulation and Dysfunction of Apoptosis. IntechOpen; 2021. doi: 10.5772/intechopen.97536.
- Chaudhry GE, Akim A, Sung YY, Tengku Muhammad TS. Cancer and apoptosis: the apoptotic activity of plant and marine natural products and their potential as targeted cancer therapeutics. Front Pharmacol 2022; 13:842376. doi: 10.3389/fphar.2022.842376 [Crossref] [ Google Scholar]
- Lim B, Greer Y, Lipkowitz S, Takebe N. Novel apoptosis-inducing agents for the treatment of cancer, a new arsenal in the toolbox. Cancers (Basel) 2019; 11(8):1087. doi: 10.3390/cancers11081087 [Crossref] [ Google Scholar]
- Pfeffer CM, Singh AT. Apoptosis: a target for anticancer therapy. Int J Mol Sci 2018; 19(2):448. doi: 10.3390/ijms19020448 [Crossref] [ Google Scholar]
- Pandey P, Lakhanpal S, Mahmood D, Kang HN, Kim B, Kang S. Bergenin, a bioactive flavonoid: advancements in the prospects of anticancer mechanism, pharmacokinetics and nanoformulations. Front Pharmacol 2024; 15:1481587. doi: 10.3389/fphar.2024.1481587 [Crossref] [ Google Scholar]
- Zhang W, Chai R. Bergenin alleviates oxidative stress and apoptosis of cardiomyocytes induced by oxygen-glucose deprivation via activating SIRT1/FOXO3a/MnSOD pathway. Curr Top Nutraceutical Res 2022; 20(3):447-53. doi: 10.37290/ctnr2641-452X.20:447-453 [Crossref] [ Google Scholar]
- Liang C, Pei S, Ju W, Jia M, Tian D, Tang Y. Synthesis and in vitro and in vivo antitumour activity study of 11-hydroxyl esterified bergenin/cinnamic acid hybrids. Eur J Med Chem 2017; 133:319-28. doi: 10.1016/j.ejmech.2017.03.053 [Crossref] [ Google Scholar]
- Pistritto G, Trisciuoglio D, Ceci C, Garufi A, D’Orazi G. Apoptosis as anticancer mechanism: function and dysfunction of its modulators and targeted therapeutic strategies. Aging (Albany NY) 2016; 8(4):603-19. doi: 10.18632/aging.100934 [Crossref] [ Google Scholar]
- Galluzzi L, Linkermann A, Kepp O, Kroemer G. Pathophysiology of cancer cell death. In: Niederhuber JE, Armitage JO, Kastan MB, Doroshow JH, Tepper JE, eds. Abeloff’s Clinical Oncology. 6th ed. Philadelphia: Elsevier; 2020. p. 74-83.e4. doi: 10.1016/b978-0-323-47674-4.00005-0.
- Jan R, Chaudhry GE. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Adv Pharm Bull 2019; 9(2):205-18. doi: 10.15171/apb.2019.024 [Crossref] [ Google Scholar]
- O’Brien MA, Kirby R. Apoptosis: a review of pro‐apoptotic and anti‐apoptotic pathways and dysregulation in disease. J Vet Emerg Crit Care 2008; 18(6):572-85. doi: 10.1111/j.1476-4431.2008.00363.x [Crossref] [ Google Scholar]
- Gano CA, Fatima S, Failes TW, Arndt GM, Sajinovic M, Mahns D. Anti-cancer potential of synergistic phytochemical combinations is influenced by the genetic profile of prostate cancer cell lines. Front Nutr 2023; 10:1119274. doi: 10.3389/fnut.2023.1119274 [Crossref] [ Google Scholar]
- Hakeem AN, El-Kersh DM, Hammam O, Elhosseiny A, Zaki A, Kamel K. Piperine enhances doxorubicin sensitivity in triple-negative breast cancer by targeting the PI3K/Akt/mTOR pathway and cancer stem cells. Sci Rep 2024; 14(1):18181. doi: 10.1038/s41598-024-65508-0 [Crossref] [ Google Scholar]
- Talib WH, Awajan D, Hamed RA, Azzam AO, Mahmod AI, Al-Yasari IH. Combination anticancer therapies using selected phytochemicals. Molecules 2022; 27(17):5452. doi: 10.3390/molecules27175452 [Crossref] [ Google Scholar]
- Kabel AM, El-Rashidy MA, Omar MS. Ameliorative potential of tamoxifen/thymoquinone combination in patients with breast cancer: a biochemical and immunohistochemical study. J Can Sci Res 2016; 1(1):1000102. doi: 10.4172/2576-1447.1000102 [Crossref] [ Google Scholar]
- Burnett JP, Lim G, Li Y, Shah RB, Lim R, Paholak HJ. Sulforaphane enhances the anticancer activity of taxanes against triple negative breast cancer by killing cancer stem cells. Cancer Lett 2017; 394:52-64. doi: 10.1016/j.canlet.2017.02.023 [Crossref] [ Google Scholar]
- Li Q, Chen S, Wang X, Cai J, Huang H, Tang S. Cisplatin-based combination therapy for enhanced cancer treatment. Curr Drug Targets 2024; 25(7):473-91. doi: 10.2174/0113894501294182240401060343 [Crossref] [ Google Scholar]